

Introduction
Dance is one of the performing arts that uses the dynamics of human movement as a form of expression, and often associates this movement with an aesthetic value. This essay will look more closely at how humans perceive dance, focusing on two important components inextricably intertwined with the world of dance. Firstly, is the idea of the dancer who has developed an extraordinary expertise in the execution of actions. Motor abilities can change not only the way a dancer performs movements, but also how s/he sees movements in general. Second, there is the implicit aesthetic response engendered in any observer while observing a dance, regardless of any previous experience or exposure to it.
Dance often involves the integration of several components (e.g. movements, costumes, music, scenography), and each of these may have artistic aspects in their own right. But the experience of a dance performance is more than the mere sum of these components; it is a complicated interaction at different levels of all components. This can render studies of dance difficult, and in order to use an experimental approach, one way is to decompose the dance so that the specific component of interest can be isolated into the simplest element. Here we will exclusively focus on the principal component of dance: the movement.
The neuroscience of seeing
How do we see dance? This is a complex question, but a first step towards the answer may be in describing how humans see movements in general. The neuroscience of action has studied movement execution for many years and their mechanisms are well known (for a review see Jeannerod, 1997; Berthoz, 2002). Besides, in the last decade a growing number of studies have investigated the neuro-cognitive mechanisms of action observation.
These studies have shown that we recruit parts of our own motor representations when we watch others’ actions (Rizzolatti & Craighero 2004). This had led to the proposal of the so-called ‘motor simulation theory’ for action perception. This theory found direct evidence for the first time in the early nineties, in electrophysiological studies performed in the laboratory of Giacomo Rizzolatti in Parma (Italy). Together with his team of neurophysiologists, Rizzolatti recorded neural responses from several regions in the primate monkey brain, while monkeys were observing different types of actions and movements.
More specifically, they recorded activity of neurons in the premotor and parietal cortices and observed how these neurons (well known for their motor properties during action execution) responded in a similar way during the mere observation of those movements that these neurons represented (di Pellegrino et al., 1992; Gallese et al., 1996). This population of neurons was called mirror neurons as they seemed to reflect, like a mirror, the observed action onto the motor representations of the observer. Their initial studies were followed by various control studies that ultimately lead to the (now widely accepted) conclusion that there is a subset of neurons in the premotor and parietal cortex that respond in a similar way to both action observation and action execution. These seminal findings could represent a first step in understanding how the human mind understands actions performed by others.
Subsequently, using modern neuroimaging techniques researchers from different laboratories have investigated in parallel whether humans might have a similar system to match observed and executed actions. These studies identified a network of areas analogous to the ones discovered in monkeys; namely in the premotor and parietal cortex, that participate both in action execution and action observation (Grafton et al. 1996; Grèzes & Decety, 2001; Buccino et al., 2001) (for a complete review, see Rizzolatti & Craighero, 2004).
These early neuroimaging studies in humans have helped to establish that humans have a similar ‘mirror system’ to the one described in non-human primates. In this way, similar areas of the human brain participate when we perform a movement, but also when we see someone else performing the same movement. The responses in these regions may reflect an internal common coding for action and perception that ultimately facilitate responses during action observation.
The discovery of the mirror neuron system supports a long-standing classical theory among philosophers and psychologists, often coined ‘motor simulation’. The general concept of the motor simulation theory for action observation is sustained by the idea that through activating the motor representation of the observed movement, I can retrieve all the information related to that action. This information is distributed in the brain, and ranges from the specific motor commands necessary to perform the movement to the general knowledge related to it (name of the action, meaning, emotions that are produced, etc.). The internal simulation of a movement may have an important role in the way we understand other people’s actions, make predictions about their movements, or read their intentions.
Dancers seeing dance
One important concept for studies of dance is ‘motor repertoire’. Motor repertoire can be defined as storage of all the motor knowledge we have acquired during our lifetime. Motor representation is the individual unit of this repertoire. The representation of each action contains the required information related to how to perform an action, including its associated sensory consequences and the meaning of that action. There are two relevant factors that constrain the human motor repertoire.
First, our movements are limited by the physical properties of the musculoskeletal system, such as the number of flexions and extensions that our joints and muscles allow (e.g. each joint allows a limited number of degrees of freedom). The second is related to motor learning. Life allows us to become motorically unique, by shaping the content of our motor repertoire when acquiring specific motor skills through a particular motor training.
This transformation is brought to perfection in dancers. Most dancers have undergone specific training in a particular dance discipline. In this way, they have shaped the content of their motor repertoire by adding the motor knowledge about these movements, allowing them to fluidly perform movements that most of us would find impossible to achieve.
According to motor simulation theory for action observation, when we watch an action we simulate it internally, by recruiting the same representations we use when we perform the action ourselves. This could provide a plausible explanation if everybody had the same motor knowledge about all action we observe. However dancers are the perfect example to illustrate that this is not necessarily the case.
This is best illustrated by one of our recent studies in which we used neuroimaging techniques to understand how motor expertise changes the way dancers (i.e. motor experts) see movements. We compared brain activity when dancers watched two different types of dance movements. Some were familiar to the dancers (familiarity was measured by how much motor training they had received in those particular dance movements); others were visually similar but dancers had actually never trained in these movements.
In order to do this, we approached two types of dancers trained in two completely different disciplines: classical ballet dancers and capoeristas. Ballet dancers will have acquired the specific motor representation of all the dance movements included in the canon of classical ballet. However they will have had little or no experience in performing or even seeing capoeira movements. The opposite applies to capoeira dancers.
We tested 10 professional ballet dancers and nine professional capoeristas from different nationalities. All participants were right-handed and aged 18 – 28. We employed functional magnetic resonance imaging (fMRI), a non-invasive technique to record brain activity. This allows for measuring which brain regions are activated when these dancers observe familiar and unfamiliar dance movements. The results of this study showed that brain activity in motor simulation regions, such as premotor and parietal cortices, were stronger when dancers watched the movements that they knew, compared to those that they had not previously performed before.
For example, we compared brain activity when ballet dancers watched ballet moves, compared to when they watched capoeira moves, and vice versa for the capoeristas (for further details, see Calvo-Merino et al., 2005). This result and subsequent studies using the present motor expertise approach (Calvo-Merino et al., 2006; Cross et al., 2006) suggest that when we see familiar movements, we recruit the information associated to that action in the same way as when we execute the movement ourselves. We therefore recover memories associated to the observed action, but only if we have previously acquired them.
The discovery of the mirror system and the idea of internal motor simulation during observation started a new line of research focused on how this mechanism can help to understand other people’s actions and intentions. A metaphor often used to illustrate this idea is that we understand others by putting ourselves ‘into their shoes’. Mirror neurons might be one of the first evidence that this idea might indeed be true. In the specific case of dance, we have shown differences during brain activity of dancers while watching familiar and non-familiar dance moves (which suggest differences during the internal motor resonance).
This internal simulation effect might be the basis of a more general ‘expertise’ mechanism that dancers use to not only perform movements with a delightful precision, but also to be able to extract much more information related to the observed move than just through mere observation. This access to the whole representation network might facilitate other important processes such as the learning of similar movements, understanding, imitation and memory for actions.
Aesthetic theories
Art is intrinsically related to aesthetics. In general, although not exclusively, art is created in order to produce an aesthetic response in an observer. At the same time, when people walk into an art gallery, or a dance theatre, they expose themselves to an aesthetic mood, to eventually reach a psychological state known as aesthetic experience (Cupchik & Laszo, 1992). Aesthetic experience has been defined as "the gratification of the sense or sensuous delights" (Goldman, 2001).
It is considered a quality uniquely reserved to the human mind, and has attracted the interest of psychologists for many years. Philosophers and psychologists have described at least two theories that have influenced studies of aesthetic experience: the objective theory and the subjective theory. The objective theory suggests that there are some particular properties of stimuli (e.g. paintings, objects, music, dance movements) that facilitate the aesthetic response. Researchers have tried to identify and describe these properties, by comparing preferences for different compositions, types of balance and symmetries (McManus & Weatherby, 1997; McManus 1980).
One such example is the ‘golden mean’ that will induce an aesthetic feeling in any observer and be preferred to any other composition of stimuli (Livio, 2002). On the other hand, subjectivist theories give larger importance to individuals’ preference, taste and attitudes. They support the common saying that "beauty is in the eye of the beholder". Studies following this perspective show a strong correlation between the level of familiarity with stimuli and the degree of likeness (Zajonc, 1968).
Nowadays, synthetic approaches are gaining ground, in which the results of a final aesthetic experience will emerge through a combination and interaction between different levels of the physical properties of external stimuli we are seeing, and the individual experience, knowledge and preference that each of us has developed during our lifetime.
With the advance of neuroscience and modern neuroimaging techniques such as fMRI or magnetoencephalography (MEG), a new discipline of so-called neuroaesthetics has focused on investigating the neural mechanisms of aesthetic processing (such as aesthetic perception and aesthetic evaluation). These studies have identified at least three types of networks or processes that participate in aesthetic experience. These are a perceptual, a cognitive and an emotional mechanism.
The perceptual mechanism is based on sensory and attentional regions (Kawabata & Zeki, 2004; Vartanian & Goel, 2004). The cognitive and emotional mechanisms are centered around the prefrontal cortex. This includes the orbitofrontal cortex, which often reflects the reward value of a stimulus, and the prefrontal dorsolateral cortex, which plays a role in interfacing perception and action and is critical for the monitoring and comparison of multiple events in working memory (Cela-Conde et al., 2004; Kawabata & Zeki, 2004; Petrides, 2000; Rolls, 2000; Vartanian & Goel, 2004).
Participants in many of these studies viewed several paintings, and then gave a rating of how much they like each of these. This approach can reveal the neural correlates of liking or disliking aesthetically-relevant stimuli. However, it provides little information about the physical properties responsible for the aesthetic experience.
In other words, one cannot project their conclusions back into a stimulus space, to identify which parameters of a stimulus lead to it being liked or disliked. This limitation is a consequence of comparing different sets of stimuli according to the attitudes expressed by each subject (Cela-Conde et al., 2004; Kawabata & Zeki, 2004).
Neural correlates of aesthetic perception of dance
Here, we present a study that aimed to go one step further, by including two new important components to our experimental conditions. The first one concerns the type of stimulus used in previous research. Previous studies have focussed on static stimuli, such as paintings. In the artistic world of performing arts, dance has a very important role. Therefore, movement seems a critical component to study when trying to understand aesthetic experience.
Second, aesthetic experience has been studied as an explicit process. This means that people were asked to rate the beauty of paintings, or to decide which painting they preferred. Our view is that the human mind can have aesthetic experience in the absence of explicit context. Therefore, taking these two issues into consideration we designed a study to investigate the neural correlates of implicit aesthetic responses to dance (Calvo-Merino et al., 2008).
The aim of this study was twofold. On one hand, it has been accepted by visual scientists that the way external stimuli are perceived depends on the intrinsic structure and properties of the visual system (Zeki & Lamb, 1994). Neuroscientists have a good idea, thanks to studies on action observation (see previous section), about how the human eye sees movements. We have provided some insight into how this also applies to dance movements.
However, we aimed to investigate how the brain regions that participate in observing dance movements are involved in representing the degree of implicit aesthetic experience an observer has while watching these movements. A second aim was to provide a physical description of the dance movements that elicited stronger or weaker responses at both the subjective level (implicit liking) and the neuronal level (how the aesthetically relevant brain regions respond to these movements).
The experimental session was carried out in two parts. In a first session, we measured brain responses in naïve participants with no formal dance experience while they watched dance movements. To ensure that participants paid attention to the movements, we asked them to rate how tiring each movements was, via keypress, on a scale from 1 – 3.
It is important to note that no explicit aesthetic question was asked during the brain scan. A range of 24 dance movements from different cultural backgrounds (classical ballet and capoeira) was selected with the help of choreographer and former classical ballet dancer Tom Sapsford to evaluate general responses to dance perception (irrespective of style).
The selected movements were classified on the bases of four kinematic properties: speed, body part used, direction of movement, and vertical and horizontal displacement. In a second session, participants rated the complete set of movements on different dimensions related to aesthetic experience (liking-disliking, simplex-complex, interesting-dull, tense-relax, weak-powerful) (Berlyne, 1974).
Although all dimensions from this questionnaire are related to aesthetic experience, only the ‘liking-disliking’ dimension showed a significant correlation with brain activity while watching dance movements. Probably because it directly reflects the personal experience of the observer, we focused on this dimension. The ratings for each movement were first normalised within each subject and then averaged across subjects to create a consensus rating of the group of participants for each movement.
Finally, the movements were divided into two categories: one contained the most-liked movements (i.e., the moves with the highest scores), the other category contained the movements with the lowest scores (more disliking). We then used this group average of all subjects’ ratings to identify brain areas sensitive to whether they were watching a generally high or low rating in this aesthetic dimension, as determined by the consensus scores (for further methodological details, see Calvo-Merino et al. 2008).
The analysis of the data produced two types of results. At the neural level, we found two specific brain regions showing significant neuroaesthetic tuning. These regions were more activated when subjects viewed movements that (on average, in the consensus) they liked, compared to movements that, on average, they disliked. We identify aesthetic sensitive areas in the visual cortex and in the premotor cortex. Visual regions participate in the early perceptual processing of the stimuli; the premotor cortex is part of a sensorimotor mechanism that links perception to action knowledge.
These two regions may be relevant for implicit positive aesthetic experience of dance. No significant results were found for the opposite comparison, i.e. when looking for brain regions sensitive to viewing less preferred rather than preferred movements. The idea of having such a sensorimotor response automatically triggered while seeing dance movements we implicitly like, may explain why dance is so easily appreciated in many human cultures.
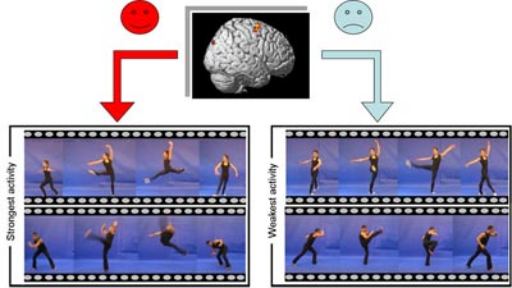 Figure 1. Sensorimotor aesthetic tuning during observation of dance movements. The top box shows a standard brain, where we have overlapped the brain responses that correlate with the group consensus aesthetic evaluation of dance movements on a scale between ‘like’ and ‘dislike’. These are premotor cortex in the right hemisphere and visual cortex bilaterally. The left panel image shows footages of the 3-second dance movements that strongly activate the premotor regions. The panel on the right illustrates the same for the weakest activity. Note that the movements in the left box (stronger activation) include horizontal and vertical displacement (jumping), while those in the left box (weaker activity) involve mainly one limb and little displacement. Video captured and edited by B. Calvo-Merino (modified from Calvo-Merino et al, 2008).
Figure 1. Sensorimotor aesthetic tuning during observation of dance movements. The top box shows a standard brain, where we have overlapped the brain responses that correlate with the group consensus aesthetic evaluation of dance movements on a scale between ‘like’ and ‘dislike’. These are premotor cortex in the right hemisphere and visual cortex bilaterally. The left panel image shows footages of the 3-second dance movements that strongly activate the premotor regions. The panel on the right illustrates the same for the weakest activity. Note that the movements in the left box (stronger activation) include horizontal and vertical displacement (jumping), while those in the left box (weaker activity) involve mainly one limb and little displacement. Video captured and edited by B. Calvo-Merino (modified from Calvo-Merino et al, 2008).At the movement level, we were able to identify stimulus quantities that specifically modulate aesthetically-relevant brain areas of the group of subjects that participated in this study. In this way, we identified which specific dance movements were responsible for maximal and minimal activation for the two aesthetically-responsive brain areas.
Figure 1 shows an example of the movements that achieve highest and lowest neural responses for the occipital area in the left hemisphere and the equivalent stimuli for the right premotor activation. Because movements were selected on the basics of four criteria, we can now produce a physical description of those dance movements that preferentially target these aesthetically sensitive areas. This suggested that, on average, these aesthetic sensitive areas preferred whole body movements, such as jumping in place or with a significant displacement of the entire body in space (e.g., horizontal jumps).
When we performed the same type of analysis based only on the behavioral data, we observed that the kinematic properties of the movements that received highest and lowest consensus liking score in the subjective rating showed clear correspondence with the moves that target the brain areas revealed as aesthetically relevant in the functional neuroimaging analysis.
Discussion and conclusions
Dance is a performing art that can elicit aesthetic experiences in the observing audience by means of actions. The neuroscientific basis of this cultural and expressive art form has been relatively little studied. The present work has explored one of many different ways of seeing dance: aesthetic perception. In particular, we have taken advantage of modern neuroimaging techniques and knowledge about the neural mechanisms involved in perception of dance movements to investigate how the brain of an observer responds during the aesthetic experience associated with seeing a dance. Our preliminary results illustrate how one can now start to investigate and study the neuroaesthetics of dance.
For this, we argue that one needs to consider a central issue that relates to most dances: a dancer that performs in front of an audience. An audience is essentially a number of independent observers. Each observer may process the dance performance in their own, highly subjective way. However, humans share a basic common neural mechanism comparable among individuals. By treating the perception of dance in the reductionist manner presented here, we were able to describe common elements during perception of dance in general, and aesthetic perception in particular. However, differences between individuals may also produce significant differences in how external stimuli are processed.
This paper presents how the motor expertise of the dancer influences the way the brain responds during dance observation. In other words, the dancer’s expertise in movement execution (motor domain) corresponds to movement perception (visual domain). In this way, observing actions that we know or are familiar with elicit stronger internal simulations of that action. One may think about this process as, if every time we watch the actions of others’, we are also ‘moving’, but subconsciously. This type of internal resonance has previously been suggested to be in the footsteps of a more complex process such as action understanding (Rizzolatti & Craighero 2004)
The (probably non-experienced) observers in the auditorium seated in front of the stage also respond to seeing the dance performance. In the studies presented here, we have focused on one type of response of the non-expert observer: the aesthetic response. We evaluated brain responses in naïve observers while they viewed dance movements, and we showed that there is a sensorimotor network that participates in the aesthetic response elicited when seeing dance. We also show that part of this network might be related with the previously described internal simulation mechanism and brain system for understanding other peoples’ actions.
However, although dancers and dance moments as a key element comprise the focus of this essay, one may question where the benefits of this type of research lie for the dance community. It may be difficult to create a partnership where both dancers and scientists profit equally. The collaboration between scientists and artists was essential in developing this project.
In the study we present here, a large part of the collaboration with the artists happened at the initial stage of the project that required the creation of the dance stimuli. While it was the experimenters’ role to define stimulus requirements to test the experimental hypotheses (e.g. kinematics properties, length, colours) it was a professional choreographer, familiar with our experimental framework and approach, who made a final selection of dance movements appropriated for the experiment, and then guided the dancers while performing the steps in the video recording session.
Valuable knowledge and interchange through interdisciplinary collaborations will be most useful in the long run, not single projects. Science often aims to understand a phenomenon (here a dance performance) by analytically deconstructing it into simpler elements, and investigating each of these elements separately. Deconstructing the whole is essential to study each independent component and its contribution to a single process.
In our studies, for example, we aim to isolate the movement from other aspects of the performance. We present small videos of movements in which dancers wear only plain black clothes and perform the selected movements in front of a blue background. However, reducing the complexity of dance to a single set of 24 movements and two performers is a grave simplification. Much more work is needed before we can start to fully combine in an experimental setting all the elements that ultimately form a dance performance.
Two approaches may be more fruitful. The one presented in this paper controls the infinite variables affecting the human mind, and focuses on one key component of the performance: the movement. A complementary approach would open room for more elements and consider simultaneously more components of the performance itself. Neither of these approaches is valid independently. One lacks ecological value, while the other exceeds in experimental noise. Therefore, only the combination of these two approaches will provide the dance community with enough scientific and artistic input to be implemented in the development of new dance material.
Finally, we showed how different movements elicited liking responses and neural responses in different ways. Because the stimuli differ only in their kinematic properties, we can consider each video clip as an independent movement unit. I could easily imagine a further study that uses a larger representation of dance movements, where each specific movement or movement element could be correlated with a subjective value in an average observer, and a specific neuronal response in aesthetic sensitive areas. In this way, choreographers could use this information to choreograph a new performance that specifically targets individual aesthetic feelings in the observer’s brain.
Therefore, our results (and future studies using a similar approach) give rise to the possibility of a ‘menu’ of dance moves, from which artists could choose those, which target aesthetically sensitive areas. In this regard, our results provide a first demonstration about the neural underpinnings of aesthetic perception in the context of the performing arts. However, this approach does not aim to imply that the creativity of the artist can be substituted or replaced by methodological implementation of this type of scientific approach. It rather suggests that choreographers can include this new knowledge as an extra tool, jointly with other creativity processes that will ultimately form the final artistic creation.
Modern neuroimaging techniques can now link the mind to the neuronal level, and the complexity of the information can be as overloading as overwhelming. This will allow the intrepid artist to reach deeper into the observer’s mind and explore further. Nevertheless, dancers and choreographers have the final word in deciding how to use the experimental approach we present here, and how to continue interactions with scientists.
Acknowledgements
This essay was part of a larger set of studies performed at University College of London with Prof. Patrick Haggard. We are grateful to Deborah Bull of the Royal Opera House (ROH2), Emma Maguire of the Royal Ballet and Giuseppe Vitolo (Professor Polvo-Capoeira Abolicao) for assistance with preparing the stimuli, and to Tom Sapsford for choreography. We thank Daniel Glaser, Julie Grèzes, Corinne Jola and Dick Passingham for help with planning and analysing the imaging studies. We thank Sven Bestmann for comments on earlier versions of the manuscript. This work was supported by a Leverhulme Trust Research Grant, Economical Social Research Council (ESRC) and City University Fellowship.
References
- Berlyne, D.E. (1974). Studies in the new experimental aesthetics: Steps toward an objective psychology of aesthetic appreciation. Washington, DC: Hemisphere Publishing.
- Berthoz, A. (2002). The Brain’s Sense of Movement. Cambridge, USA: Harvard University Press.
- Buccino, G., Binkofski, F., Fink, G. R., Fadiga, L., Fogassi, L. et al., (2001). Action observation activates premotor and parietal areas in a somatotopic manner: an fMRI study. Eur J Neurosi, 13, 400 – 404.
- Calvo-Merino, B., Glaser, D. E., Grèzes, J., Passingham, R. E., & Haggard, P. (2005). Action observation and acquired motor skills: An FMRI study with expert dancers. Cerebral Cortex, 15, 1243 – 1249.
- Calvo-Merino, B., Grèzes, J., Glaser, D. E., Passingham, R. E., & Haggard, P. (2006). Seeing or doing? Influence of visual and motor familiarity in action observation. Current Biology, 16, 1905 – 1910.
- Calvo-Merino, B., Jola, C., Glaser, D. E., & Haggard, P. (2008). Towards a sensorimotor aesthetics of performing art. Consciousness and Cognition, 17, 911 – 922.
- Cela-Conde, C. J., Marty, G., Maestu, F., Ortiz, T., Munar, E., Fernandez, A. et al. (2004). Activation of the prefrontal cortex in the human visual aesthetic perception. Proceedings of the National Academy of Sciences of the United States of America, 101, 6321 – 6325.
- Cross, E. S., Hamilton, A. F. D. C., & Grafton, S. T. (2006). Building a motor simulation de novo: Observation of dance by dancers. Neuroimage, 31, 1257 – 1267.
- Cupchik, G. C., & László, J. (1992). Emerging visions of the aesthetic process psychology semiology and philosophy. Cambridge, New York, USA: Cambridge University Press.
- di Pellegrino, G., Fadiga, L., Fogassi, L., Gallese, V., & Rizzolatti, G. (1992). Understanding motor events: a neurophysiological study. Experimental Brain Research, 91, 176 – 180.
- Gallese, V., Fadiga, L., Fogassi, L., & Rizzolatti, G. (1996). Action recognition in the premotor cortex. Brain, 119, 593–609.
- Grèzes, J., & Decety, J. (2001). Functional anatomy of execution, mental simulation, observation, and verb generation of actions: a metaanalysis. Human Brain Mapping, 12, 1 – 19.
- Grafton, S. T., Arbib, M. A., Fadiga, L., & Rizzolatti, G. (1996). Localisation of grasp representations in humans by positron emission tomography. 2. Observation compared with imagination. Experimental Brain Research, 112, 103 – 111.
- Goldman, A. (2001). The Routledge companion to aesthetics. In B. Gaut & D. McIver Lopes (Eds.), The aesthetic (pp. 181–192). London: Routledge.
- Jeannerod, M. (1997). The cognitive neuroscience of action. Oxford: Blackwell.
- Kawabata, H., & Zeki, S. (2004). Neural correlates of beauty. Journal of Neurophysiology, 91, 1699 – 1705.
- Livio, M. (2002). The Golden Ratio. The story of Phi, the extraordinary number of nature, art and beauty. London: Headline Book Publishing.
- McManus, I. C. (1980). The aesthetics of simple figures. British Journal of Psychology, 71, 505 – 524.
- McManus, I. C., & Weatherby, P. (1997). The golden section and the aesthetics of form and composition: A cognitive model. Empirical Studies of the Arts, 15(2), 209 – 232.
- Petrides, M. (2000). The role of the mid-dorsolateral prefrontal cortex in working memory. Experimental Brain Research, 133, 44 – 54.
- Rizzolatti, G., & Craighero, L. (2004). The mirror-neuron system. Annual Review of Neuroscience, 27, 169 – 92.
- Rolls, E.T. (2000). Precis of the brain and emotion. The Behavioral and Brain Sciences, 23, 177 – 191.
- Vartanian, O., & Goel, V. (2004). Neuroanatomical correlates of aesthetic preference for paintings. Neuroreport, 15, 893 – 897.
- Zajonc, R. B. (1968). Attitudinal effects of mere exposure. Journal of Personality and Social Psychology, 9, 1 – 27.
- Zeki, S., & Lamb, M. (1994). The neurology of kinetic art. Brain, 117, 607 – 636.